
 Neurons and Synapses
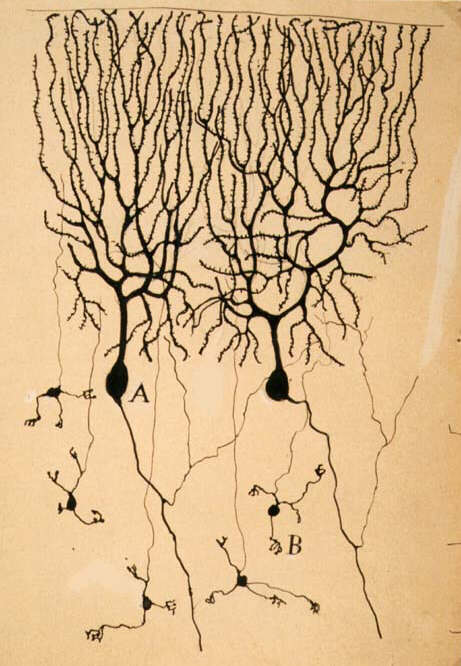
Neurons and SynapsesHaving established that neurons do not fuse together at any level of their ramifications, a nagging research question became important. How is the contact between neurons? How the transmission of an electrical impulse wave occurs between two neurons?
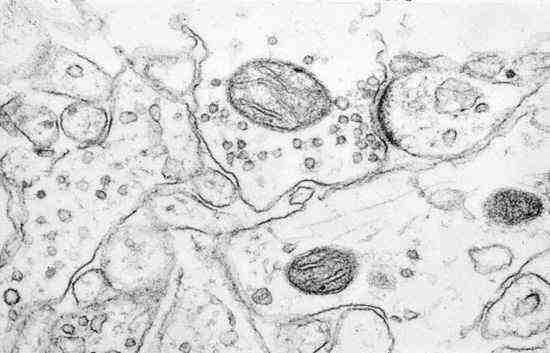
This "neuronal junction", how it was called then, was too small to be seen by the microscopes of that time, and the "gap" which surely occurs at the point of contact could not be documented. In fact, the definitive morphological proof for this came only in 1954, with the work of George Palade, Eduardo de Robertis and George Bennett, who used the recently invented electron microscope, which provided the higher magnifications to see the synaptic ultrastructure. They showed the existence of distinct presynaptic and postsynaptic elements, a synaptic cleft and presynaptic vesicles.
The problem of the nature of transmission from one neuron to another was also a point of major consideration and inquiry amongst neurophysiologists at the turn of the 20th century. Many defended the idea that the transmission should be electrical, just like the propagation wave along the axon. Back in 1846, Emil DuBois-Reymond proposed the existence of synapses and that they could be either electrical or chemical. He had no support for this speculation, and so it went into oblivion. It made great sense to imagine electrical synapses (actually, some scientists even imagined little electrical sparks crossing the synaptic gap!), because it yielded a simpler picture for the nervous system. Unfortunately there were three important pieces of evidence that ran against it.
Many of the experiments which provided these data were carried out in the laboratory of the great British physiologist Sir Charles S. Sherrington (1852-1952), who investigated in the late 1890s the physiology of simple and complex motor reflexes. His brilliant work led to the concept of the integrative action of the nervous system, complementing the line of reasoning started by the "father" of physiology, Claude Bernard (1813-1878), in the previous century. Sherrington argued that the junction between neurons was the final pathway of regulation of transmission in the nervous system, and gave it a name, "synapse", which, in Greek, means "to clasp".
|
|
|
|
Sherrington´s work (who also got a Nobel Prize, in 1921), highlighted a number of properties of reflex arches at the spinal level, and how they are influenced and modulated by brain structures at a higher level, such as the cerebellum and the brainstem. He was one of the first to discover that the interplay of central excitation and inhibition are fundamental for this integration, and, therefore, that synapses should be at least of these two types. This was a new functional concept. His work was immortalized in one of the most classical texts of modern neurophysiology, "The Integrative Action of the Nervous System", first published in 1906.
The same concepts of excitation and inhibition appeared again in the study of the peripheral nervous system, particularly in the Autonomous Nervous System (ANS). This is the part of the nervous system which is responsible for the control over visceral organs, such as the heart, the blood vessels, the gastrointestinal tube and glands, the urinary system, and so forth. Since the 19th century, neurophysiologists and neuroanatomists were able to study how the ANS distributed throughout the organism and what were its basic actions.
For example, they discovered that the force of contraction of heart muscles (which is also an striated muscle) and the heart rate (frequency of beating) could be altered in two opposite directions by two different subdivisions of the ANS. Force of contraction and heart rate increases when the so-called sympathetic division is activated (by electrically stimulating the ganglionar nerves which innervate the heart, for example). They are decreased when the vagus nerve, which conduits the so-called parasympathetic division, is activated.
Since the target organs for these two nerve systems are the same, therefore one must logically conclude that there must be two different actions at their synapses with heart muscle: one excitatory, another one inhibitory. The great mistery, therefore, was to imagine how electrical transmission in the same postsynaptic element would achieve totally opposite effects. The solution, of course, was again a chemical transmission. A strong evidence for this would be the discovery of active substances that could reproduce the effects of ANS activation at some target organs.
This evidence was already considerable: for example, Henry Halett Dale (1865-1968), a British physiologist, investigated in 1914 the active substance in ergot, which was extracted from a fungal infection of wheat, which provoked strong central nervous system effects, including death. By accident, he made the discovery that ergot would reverse the effects of adrenaline. He was able to study the effects of this substance on the heart and found that they were identical to the actions of the parasympathetic ANS. This substance was later chemically identified as being acetylcholine. In another line of work, British scientist T.R. Elliott had discovered in 1904 that the extract of the medulla of the adrenal gland (with a substance called adrenaline) was shown to mimick almost exactly the actions of the sympathetic nervous system.
All these evidences convinced neuroscientists in the beginning of the 20th century that most synapses had chemical transmission. But the fundamental proof for this came only in 1921, as we will see, with the crucial experiments carried out by Otto Loewi.
|
Neurons and Synapses: The History of Its Discovery Part 4 of 6 |
 |



{kind=link}
{kind=link}